



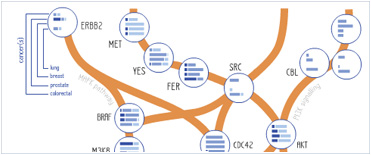







 Foundational characteristics of cancer include proliferation, angiogenesis, migration, evasion of apoptosis, and cellular immortality. Find key markers for these cellular processes and antibodies to detect them.
Foundational characteristics of cancer include proliferation, angiogenesis, migration, evasion of apoptosis, and cellular immortality. Find key markers for these cellular processes and antibodies to detect them. The SUMOplot™ Analysis Program predicts and scores sumoylation sites in your protein. SUMOylation is a post-translational modification involved in various cellular processes, such as nuclear-cytosolic transport, transcriptional regulation, apoptosis, protein stability, response to stress, and progression through the cell cycle.
The SUMOplot™ Analysis Program predicts and scores sumoylation sites in your protein. SUMOylation is a post-translational modification involved in various cellular processes, such as nuclear-cytosolic transport, transcriptional regulation, apoptosis, protein stability, response to stress, and progression through the cell cycle. The Autophagy Receptor Motif Plotter predicts and scores autophagy receptor binding sites in your protein. Identifying proteins connected to this pathway is critical to understanding the role of autophagy in physiological as well as pathological processes such as development, differentiation, neurodegenerative diseases, stress, infection, and cancer.
The Autophagy Receptor Motif Plotter predicts and scores autophagy receptor binding sites in your protein. Identifying proteins connected to this pathway is critical to understanding the role of autophagy in physiological as well as pathological processes such as development, differentiation, neurodegenerative diseases, stress, infection, and cancer.
PINK1 (PARK6) Antibody (N-term T133)
Purified Rabbit Polyclonal Antibody (Pab)


- SPECIFICATION
- CITATIONS: 2
- PROTOCOLS
- BACKGROUND

Application
| IHC-P, WB, E |
|---|---|
| Primary Accession | Q9BXM7 |
| Other Accession | NP_115785 |
| Reactivity | Human |
| Host | Rabbit |
| Clonality | Polyclonal |
| Isotype | Rabbit IgG |
| Calculated MW | 62769 Da |
| Antigen Region | 118-147 aa |
| Gene ID | 65018 |
|---|---|
| Other Names | Serine/threonine-protein kinase PINK1, mitochondrial, BRPK, PTEN-induced putative kinase protein 1, PINK1 |
| Target/Specificity | This PINK1 (PARK6) antibody is generated from rabbits immunized with a KLH conjugated synthetic peptide between 118-147 amino acids from the N-terminal region of human PINK1 (PARK6). |
| Dilution | WB~~1:1000 IHC-P~~1:50~100 |
| Format | Purified polyclonal antibody supplied in PBS with 0.09% (W/V) sodium azide. This antibody is prepared by Saturated Ammonium Sulfate (SAS) precipitation followed by dialysis against PBS. |
| Storage | Maintain refrigerated at 2-8°C for up to 2 weeks. For long term storage store at -20°C in small aliquots to prevent freeze-thaw cycles. |
| Precautions | PINK1 (PARK6) Antibody (N-term T133) is for research use only and not for use in diagnostic or therapeutic procedures. |
| Name | PINK1 |
|---|---|
| Function | Serine/threonine-protein kinase which acts as a sensor of mitochondrial damage and protects against mitochondrial dysfunction during cellular stress. It phosphorylates mitochondrial proteins to coordinate mitochondrial quality control mechanisms that remove and replace dysfunctional mitochondrial components (PubMed:14607334, PubMed:15087508, PubMed:18443288, PubMed:18957282, PubMed:19229105, PubMed:19966284, PubMed:20404107, PubMed:20547144, PubMed:20798600, PubMed:22396657, PubMed:23620051, PubMed:23754282, PubMed:23933751, PubMed:24660806, PubMed:24751536, PubMed:24784582, PubMed:24896179, PubMed:24898855, PubMed:25527291, PubMed:32484300). Depending on the severity of mitochondrial damage, activity ranges from preventing apoptosis and stimulating mitochondrial biogenesis to eliminating severely damaged mitochondria via PINK1-PRKN-dependent mitophagy (PubMed:14607334, PubMed:15087508, PubMed:18443288, PubMed:19966284, PubMed:20404107, PubMed:20798600, PubMed:22396657, PubMed:23620051, PubMed:23933751, PubMed:24898855, PubMed:32047033, PubMed:32484300). When cellular stress results in irreversible mitochondrial damage, PINK1 accumulates at the outer mitochondrial membrane (OMM) where it phosphorylates pre-existing polyubiquitin chains at 'Ser-65', recruits PRKN from the cytosol to the OMM and activates PRKN by phosphorylation at 'Ser-65'; activated PRKN then ubiquinates VDAC1 and other OMM proteins to initiate mitophagy (PubMed:14607334, PubMed:15087508, PubMed:19966284, PubMed:20404107, PubMed:20798600, PubMed:23754282, PubMed:23933751, PubMed:24660806, PubMed:24751536, PubMed:24784582, PubMed:25474007, PubMed:25527291, PubMed:32047033). The PINK1-PRKN pathway also promotes fission of damaged mitochondria through phosphorylation and PRKN-dependent degradation of mitochondrial proteins involved in fission such as MFN2 (PubMed:18443288, PubMed:23620051, PubMed:24898855). This prevents the refusion of unhealthy mitochondria with the mitochondrial network or initiates mitochondrial fragmentation facilitating their later engulfment by autophagosomes (PubMed:18443288, PubMed:23620051). Also promotes mitochondrial fission independently of PRKN and ATG7-mediated mitophagy, via the phosphorylation and activation of DNM1L (PubMed:18443288, PubMed:32484300). Regulates motility of damaged mitochondria by promoting the ubiquitination and subsequent degradation of MIRO1 and MIRO2; in motor neurons, this likely inhibits mitochondrial intracellular anterograde transport along the axons which probably increases the chance of the mitochondria undergoing mitophagy in the soma (PubMed:22396657). Required for ubiquinone reduction by mitochondrial complex I by mediating phosphorylation of complex I subunit NDUFA10 (By similarity). Phosphorylates LETM1, positively regulating its mitochondrial calcium transport activity (PubMed:29123128). |
| Cellular Location | Mitochondrion outer membrane; Single-pass membrane protein. Mitochondrion inner membrane {ECO:0000250|UniProtKB:Q99MQ3}; Single-pass membrane protein. Cytoplasm, cytosol. Note=Localizes mostly in mitochondrion and the two smaller proteolytic processed fragments localize mainly in cytosol (PubMed:19229105). Upon mitochondrial membrane depolarization following damage, PINK1 import into the mitochondria is arrested, which induces its accumulation in the outer mitochondrial membrane, where it acquires kinase activity (PubMed:18957282) |
| Tissue Location | Highly expressed in heart, skeletal muscle and testis, and at lower levels in brain, placenta, liver, kidney, pancreas, prostate, ovary and small intestine. Present in the embryonic testis from an early stage of development |

Provided below are standard protocols that you may find useful for product applications.
Background
Defects in PINK1 are the cause of autosomal recessive early-onset Parkinson's disease 6 (PARK6). Six novel pathogenic PINK1 mutations suggest that PINK1 may be the second most common causative gene next to parkin in parkinsonism with the recessive mode of inheritance. Strong evidence indicates that, although important in mendelian forms of Parkinson's disease (PD), PINK1 does not influence the cause of sporadic nonmendelian forms of PD.
References
Rogaeva,E.,et al. Arch. Neurol. 61 (12), 1898-1904 (2004)
Hatano,Y., et al. Ann. Neurol. 56 (3), 424-427 (2004)
Healy,D.G., et al. Ann. Neurol. 56 (3), 329-335 (2004)
Valente,E.M., et al. Science 304 (5674), 1158-1160 (2004)
Nakajima,A., et al. Cancer Lett. 201 (2), 195-201 (2003)
Unoki,M. and Nakamura,Y. Oncogene 20 (33), 4457-4465 (2001)
If you have used an Abcepta product and would like to share how it has performed, please click on the "Submit Review" button and provide the requested information. Our staff will examine and post your review and contact you if needed.
If you have any additional inquiries please email technical services at tech@abcepta.com.
Ordering Information
Other Products
Shipping Information
