


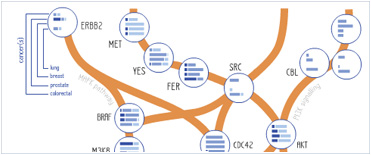







 Foundational characteristics of cancer include proliferation, angiogenesis, migration, evasion of apoptosis, and cellular immortality. Find key markers for these cellular processes and antibodies to detect them.
Foundational characteristics of cancer include proliferation, angiogenesis, migration, evasion of apoptosis, and cellular immortality. Find key markers for these cellular processes and antibodies to detect them. The SUMOplot™ Analysis Program predicts and scores sumoylation sites in your protein. SUMOylation is a post-translational modification involved in various cellular processes, such as nuclear-cytosolic transport, transcriptional regulation, apoptosis, protein stability, response to stress, and progression through the cell cycle.
The SUMOplot™ Analysis Program predicts and scores sumoylation sites in your protein. SUMOylation is a post-translational modification involved in various cellular processes, such as nuclear-cytosolic transport, transcriptional regulation, apoptosis, protein stability, response to stress, and progression through the cell cycle. The Autophagy Receptor Motif Plotter predicts and scores autophagy receptor binding sites in your protein. Identifying proteins connected to this pathway is critical to understanding the role of autophagy in physiological as well as pathological processes such as development, differentiation, neurodegenerative diseases, stress, infection, and cancer.
The Autophagy Receptor Motif Plotter predicts and scores autophagy receptor binding sites in your protein. Identifying proteins connected to this pathway is critical to understanding the role of autophagy in physiological as well as pathological processes such as development, differentiation, neurodegenerative diseases, stress, infection, and cancer.
SFPQ Antibody (C-term)
Affinity Purified Rabbit Polyclonal Antibody (Pab)

- SPECIFICATION
- CITATIONS
- PROTOCOLS
- BACKGROUND

Application
| WB, E |
|---|---|
| Primary Accession | P23246 |
| Other Accession | Q8VIJ6, NP_005057.1 |
| Reactivity | Human |
| Predicted | Mouse |
| Host | Rabbit |
| Clonality | Polyclonal |
| Isotype | Rabbit IgG |
| Calculated MW | 76149 Da |
| Antigen Region | 676-705 aa |
| Gene ID | 6421 |
|---|---|
| Other Names | Splicing factor, proline- and glutamine-rich, 100 kDa DNA-pairing protein, hPOMp100, DNA-binding p52/p100 complex, 100 kDa subunit, Polypyrimidine tract-binding protein-associated-splicing factor, PSF, PTB-associated-splicing factor, SFPQ, PSF |
| Target/Specificity | This SFPQ antibody is generated from rabbits immunized with a KLH conjugated synthetic peptide between 676-705 amino acids from the C-terminal region of human SFPQ. |
| Dilution | WB~~1:2000 |
| Format | Purified polyclonal antibody supplied in PBS with 0.09% (W/V) sodium azide. This antibody is purified through a protein A column, followed by peptide affinity purification. |
| Storage | Maintain refrigerated at 2-8°C for up to 2 weeks. For long term storage store at -20°C in small aliquots to prevent freeze-thaw cycles. |
| Precautions | SFPQ Antibody (C-term) is for research use only and not for use in diagnostic or therapeutic procedures. |
| Name | SFPQ |
|---|---|
| Synonyms | PSF |
| Function | DNA- and RNA binding protein, involved in several nuclear processes. Essential pre-mRNA splicing factor required early in spliceosome formation and for splicing catalytic step II, probably as a heteromer with NONO. Binds to pre-mRNA in spliceosome C complex, and specifically binds to intronic polypyrimidine tracts. Involved in regulation of signal-induced alternative splicing. During splicing of PTPRC/CD45, a phosphorylated form is sequestered by THRAP3 from the pre-mRNA in resting T-cells; T-cell activation and subsequent reduced phosphorylation is proposed to lead to release from THRAP3 allowing binding to pre-mRNA splicing regulatotry elements which represses exon inclusion. Interacts with U5 snRNA, probably by binding to a purine- rich sequence located on the 3' side of U5 snRNA stem 1b. May be involved in a pre-mRNA coupled splicing and polyadenylation process as component of a snRNP-free complex with SNRPA/U1A. The SFPQ-NONO heteromer associated with MATR3 may play a role in nuclear retention of defective RNAs. SFPQ may be involved in homologous DNA pairing; in vitro, promotes the invasion of ssDNA between a duplex DNA and produces a D-loop formation. The SFPQ-NONO heteromer may be involved in DNA unwinding by modulating the function of topoisomerase I/TOP1; in vitro, stimulates dissociation of TOP1 from DNA after cleavage and enhances its jumping between separate DNA helices. The SFPQ-NONO heteromer binds DNA (PubMed:25765647). The SFPQ-NONO heteromer may be involved in DNA non-homologous end joining (NHEJ) required for double-strand break repair and V(D)J recombination and may stabilize paired DNA ends; in vitro, the complex strongly stimulates DNA end joining, binds directly to the DNA substrates and cooperates with the Ku70/G22P1-Ku80/XRCC5 (Ku) dimer to establish a functional preligation complex. SFPQ is involved in transcriptional regulation. Functions as a transcriptional activator (PubMed:25765647). Transcriptional repression is mediated by an interaction of SFPQ with SIN3A and subsequent recruitment of histone deacetylases (HDACs). The SFPQ-NONO-NR5A1 complex binds to the CYP17 promoter and regulates basal and cAMP-dependent transcriptional activity. SFPQ isoform Long binds to the DNA binding domains (DBD) of nuclear hormone receptors, like RXRA and probably THRA, and acts as a transcriptional corepressor in absence of hormone ligands. Binds the DNA sequence 5'-CTGAGTC-3' in the insulin-like growth factor response element (IGFRE) and inhibits IGF1-stimulated transcriptional activity. Regulates the circadian clock by repressing the transcriptional activator activity of the CLOCK-BMAL1 heterodimer. Required for the transcriptional repression of circadian target genes, such as PER1, mediated by the large PER complex through histone deacetylation (By similarity). Required for the assembly of nuclear speckles (PubMed:25765647). Plays a role in the regulation of DNA virus-mediated innate immune response by assembling into the HDP-RNP complex, a complex that serves as a platform for IRF3 phosphorylation and subsequent innate immune response activation through the cGAS-STING pathway (PubMed:28712728). |
| Cellular Location | Nucleus speckle. Nucleus matrix. Cytoplasm. Note=Predominantly in nuclear matrix |

Thousands of laboratories across the world have published research that depended on the performance of antibodies from Abcepta to advance their research. Check out links to articles that cite our products in major peer-reviewed journals, organized by research category.
info@abcepta.com, and receive a free "I Love Antibodies" mug.

Provided below are standard protocols that you may find useful for product applications.
Background
DNA-and RNA binding protein, involved in several nuclear processes. Essential pre-mRNA splicing factor required early in spliceosome formation and for splicing catalytic step II, probably as an heteromer with NONO. Binds to pre-mRNA in spliceosome C complex, and specifically binds to intronic polypyrimidine tracts. Interacts with U5 snRNA, probably by binding to a purine-rich sequence located on the 3' side of U5 snRNA stem 1b. May be involved in a pre-mRNA coupled splicing and polyadenylation process as component of a snRNP-free complex with SNRPA/U1A. The SFPQ-NONO heteromer associated with MATR3 may play a role in nuclear retention of defective RNAs. SFPQ may be involved in homologous DNA pairing; in vitro, promotes the invasion of ssDNA between a duplex DNA and produces a D-loop formation. The SFPQ-NONO heteromer may be involved in DNA unwinding by modulating the function of topoisomerase I/TOP1; in vitro, stimulates dissociation of TOP1 from DNA after cleavage and enhances its jumping between separate DNA helices. The SFPQ-NONO heteromer may be involved in DNA nonhomologous end joining (NHEJ) required for double-strand break repair and V(D)J recombination and may stabilize paired DNA ends; in vitro, the complex strongly stimulates DNA end joining, binds directly to the DNA substrates and cooperates with the Ku70/G22P1-Ku80/XRCC5 (Ku) dimer to establish a functional preligation complex. SFPQ is involved in transcriptional regulation. Transcriptional repression is probably mediated by an interaction of SFPQ with SIN3A and subsequent recruitment of histone deacetylases (HDACs). The SFPQ-NONO/SF-1 complex binds to the CYP17 promoter and regulates basal and cAMP-dependent transcriptional avtivity. SFPQ isoform Long binds to the DNA binding domains (DBD) of nuclear hormone receptors, like RXRA and probably THRA, and acts as transcriptional corepressor in absence of hormone ligands. Binds the DNA sequence 5'-CTGAGTC-3' in the insulin-like growth factor response element (IGFRE) and inhibits IGF-I-stimulated transcriptional activity.
References
Heyd, F., et al. Mol. Cell 40(1):126-137(2010)
Marko, M., et al. Exp. Cell Res. 316(3):390-400(2010)
David, A., et al. Eur. J. Endocrinol. 162(1):37-42(2010)
Olsen, J.V., et al. Cell 127(3):635-648(2006)
Ong, S.E., et al. Nat. Methods 1(2):119-126(2004)
If you have used an Abcepta product and would like to share how it has performed, please click on the "Submit Review" button and provide the requested information. Our staff will examine and post your review and contact you if needed.
If you have any additional inquiries please email technical services at tech@abcepta.com.
Ordering Information
Other Products
Shipping Information
